
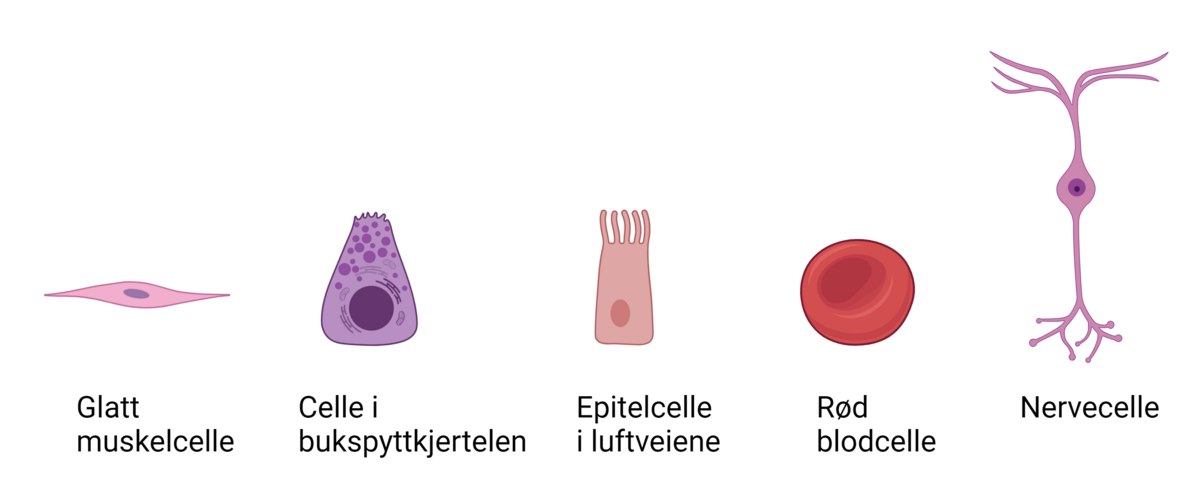
Genregulering er når cellene regulerer hvilke gener som er slått av eller på, i hvilke vev og organer de er aktive, og hvor mye genprodukt som lages til enhver tid. Genene er oppskrifter for hvordan en organisme skal se ut og fungere. Alle cellene i en flercellet organisme inneholder de samme genene, men forskjellige gener kommer til uttrykk i forskjellige deler av kroppen (vev). Ikke alle gener skal være aktive hele tiden i alle celler, derfor er genregulering nødvendig.
Alle organismer har genregulering. Mekanismene er noe forskjellig mellom prokaryote (bakterier og arkebakterier) og eukaryote organismer (sopp, planter og dyr), men noen ting er felles.




Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.